


Рыбацкие статьи
О причинах сокращения численности популяции невской корюшки
Чукучан, Рыбовод
Уважаемые господа-любители! Зовут меня Алексей Николаевич, более тридцати лет занимаюсь вопросами биологии и численности корюшки Финского залива. Захожу на Форум и у меня сложилось впечатление, что вы имеете слабое представление об объекте вашего лова - корюшке Финского залива+" http://fisher.spb.ru/forums/index.php?t=65
Этот пост специалиста рыбохозяйственной отрасли А.Н.Попова, в том числе содержащий ссылки на авторские научные публикации, послужил началом довольно горячей дискуссии, развернувшейся на форуме вокруг питерской корюшки и проблемы сокращения ее численности в последние годы. Поучаствовав в том давнем обмене мнениями и расценив некоторые выводы А.Н.Попова как достаточно сомнительные, мы, будучи профессиональными ихтиологами, решили высказать собственное суждение по данному вопросу. Свои мысли мы оформили в виде статьи, которая недавно была опубликована в журнале, издающимся под патронажем Федерального Агентства по Рыболовству.
Считаем своей приятной обязанностью поблагодарить всех форумчан, некогда принявших живое участие в корюшиной дискуссии, и дать возможность желающим ознакомиться с результатами нашего коллективного ПКРовского творчества, поскольку все вы - кто был тогда, что называется, и "за", и "против" - полноправные соавторы публикации. Без вас мы едва ли когда-нибудь взялись за это дело.
Считаем своей приятной обязанностью поблагодарить всех форумчан, некогда принявших живое участие в корюшиной дискуссии, и дать возможность желающим ознакомиться с результатами нашего коллективного ПКРовского творчества, поскольку все вы - кто был тогда, что называется, и "за", и "против" - полноправные соавторы публикации. Без вас мы едва ли когда-нибудь взялись за это дело.
Чукучан, Рыбовод
вопросы рыболовства , 2010, том 11, №3(43), с. 514-533
problems of fisheries , 2010, vol. 11, №3(43), p. 514-533
© 2010 г. Д.С. Сендек, А.Е. Королевсударственный научно-исследовательский институт озерного и речного рыбного хозяйства, Санкт-Петербург 199053Поступила в редакцию 04.02.2010 г.Окончательный вариант получен 11.06.2010 г.Статья обобщает сведения о причинах сокращения численности популяции невской корюшки в связи с изменениями экологической ситуации в Невской губе на протяжении последних десятилетий. Среди негативных факторов, обуславливающих сокращение запасов корюшки, основными являются ухудшение кормовой базы рыб в заливе, влияние дамбы КЗС, интенсификация гидротехнических работ в Невской губе, неучтенный вылов, некорректный расчет величины общего допустимого улова. Предлагается ряд конкретных мероприятий по улучшению положения популяции невской корюшки.Ключевые слова: невская корюшка; промысловые запасы; эвтрофикация; гидротехнические работы.
Биологический статус невской корюшки – полупроходная рыба, но для Петербурга и его жителей значение этой рыбы отнюдь не полу- и, тем более, не проходное: испокон веков корюшка являлась одной из самых доступных по цене и отменной по вкусовым качествам рыбой Финского залива, и поэтому по праву считается историческим символом города. Однако, в последние годы на фоне продолжающейся популяризации корюшки в качестве одного из городских брендов (проведение ставшего традиционным весеннего фестиваля корюшки у стен Петропавловской крепости) наблюдается значительное сокращение ее запасов. Снижение численности этой рыбы, относящейся к виду Osmerus eperlanus, столь значительно, что спрос на рынке Петербурга стал восполняться за счет мелкой ладожской корюшки, а также Европейской корюшки, импортируемой из соседней Прибалтики. В последние два года под видом местной корюшки в торговле стала появляться Азиатская зубатая корюшка, Osmerus mordax dentex, завозимая в замороженном виде из бассейнов Белого и Баренцева морей и с Дальнего Востока.
В отличие от более мелких, преимущественно озерных форм Европейской корюшки, обитающих в бассейнах Балтийского, Белого и Баренцева морей, полупроходная корюшка Финского залива нагуливается в открытой части залива, богатого кормом, а во время весеннего нереста подходит к берегам, где ищет пригодные для икрометания песчаные отмели. В самой восточной части Финского залива такие благоприятные для нереста условия невская корюшка находит в распресненных зонах вдоль берегов Невской губы, а также в самой реке Неве (Кожевников, 1949; Подаруева, 1972; Неелов, 1987; Попов, 2006а). Считается, что в меньшей степени корюшка может нереститься у северного побережья залива (р-н Сестрорецка), а также в реках Луга, Нарва, Систа и др. (Кудерский, 2005). Ближе к границе с Финляндией, в Выборгском заливе и прилегающих акваториях, обитает локальная популяция корюшки, которая, в отличие от невской, не совершает протяженных нагульных миграций и характеризуется невысоким темпом роста (Попов, 2006а).
Еще в недавнем прошлом площади нерестилищ в Невской губе и Неве были таковы, что в некоторые годы обеспечивали промысловый запас нерестовой корюшки порядка 250 млн. шт., при этом уловы достигали 3,5 тыс. т (Попов, 2006а).
Развивающиеся икринки и личинки невской корюшки с нерестовых площадей сносятся течением к западу, на выход из Невской губы. По данным середины 1970-1980-х годов (Галкина, Широков, 1978; Попов, 2006а), наибольшая концентрация личинок корюшки в июле обнаруживалась в районе Зеленогорска-Сестрорецка, а также недалеко от Стрельны, в районе сравнительно малоподвижных вод Южной Лахтинской отмели. По мере роста личинки из поверхностных слоев воды постепенно переходят в придонную зону, где корюшка и проводит большую часть своей взрослой жизни в заливе, которая может длиться до 9-10 лет (Кожевников, 1949).
Промысел корюшки ведется преимущественно в прибрежной части залива и в реке Нева во время нерестового хода рыбы, который приходится на апрель-май. В промысловых уловах преобладают особи 3-5 лет. Согласно данных официальной статистики, наиболее значительные уловы корюшки за последние пятьдесят лет наблюдались в 1985-1989 гг. (средний ежегодный улов – 3 056 т), в завершающее десятилетие прошлого века они сократились, соответственно, до 1996 т в год, а за период 2003-2007 гг. упали еще существеннее и не превышали 200 т в год. Наконец, по данным Севзапрыбвода, в 2008 г. был поставлен абсолютный «рекорд» последних десятилетий – поймано всего 115,7 т корюшки.
Общепризнано, что причин резкого снижения запасов невской корюшки несколько, однако даже среди специалистов-ихтиологов нет единого мнения о степени влияния того или иного фактора. Наиболее подробно негативные тенденции в динамике численности популяции корюшки рассмотрены в серии научных публикаций А.Н. Попова (Попов, 1999, 2004, 2006а). Среди основных естественных факторов среды, определяющих эффективность воспроизводства корюшки, автор статей выделяет два главных – численность родительского запаса и температуру воды в нерестовый период. Другие причины носят преимущественно антропогенный характер, и, как полагает А.Н. Попов, они начали сказываться на популяции невской корюшки в 1980-1990-е гг. Среди этих факторов автор статей рассматривает следующие:
- катастрофическое снижение численности основных объектов питания корюшки – мизид и амфипод;
- значительное сокращение уровня естественного воспроизводства корюшки;
- небывалый рост подледного любительского рыболовства.
Вывод А.Н. Попова о качественном и количественном сокращении кормовой базы корюшки на первый взгляд очевиден. Считается, что продуктивная емкость Балтийского моря в последние годы в целом заметно снизилась (Кудерский и др., 2008), и из-за выпадения из рациона нагуливающейся в Восточной части Финского залива корюшки главных кормовых объектов (придонных ракообразных – амфипод и мизид), рыба недоедает, вынуждена переходить на второстепенные объекты питания. Как следствие нехватки пищевых ресурсов, начиная с периода 1991-2001 гг. существенно замедлился темп роста корюшки, рыба стала мельчать (Попов, 2006а). Той же причиной принято объяснять ухудшение положения другой массовой рыбы залива – салаки, чьи запасы в последние годы также существенно сократились (Попов, 2006б).
В связи с резким уменьшением численности реликтовых ракообразных в более глубоких частях Балтики для всех популяций корюшки Балтийского моря, казалось бы, можно было ожидать увидеть схожую тенденцию сокращения численности. Однако данные по промысловой статистике корюшки в соседних Прибалтийских странах ставят под сомнение справедливость тезиса о жесткой зависимости благополучия популяций корюшки от обилия фауны реликтовых ракообразных.
Согласно данных финских промысловиков (официальная статистика предоставлена Finnish Game and Fisheries Research Institute), в последние двенадцать лет регистрируется постепенное снижение уловов корюшки вдоль всего побережья Финляндии (с 1 252 т в 1996 г. до 227 т в 2007 г.), однако кривая падения не столь продолжительна и не так крута, как «пике» на графике, построенном по данным о ежегодных уловах в российской части Финского залива (с 3 536 т в 1989 г. до 116 т в 2008 г.) (pис. 1).

Рис. 1. Динамика уловов корюшки в российской части Финского залива и в сопредельных странах.
Fig. 1. Dynamics of smelt catches in Russian part of the Gulf of Finland and adjacent countries.
Кроме того, минимальные финские уловы корюшки последних лет 2004-
2007 гг. (ежегодно в среднем 228 т) сопоставимы с низкими уловами финских рыбаков за период 1985-1989 гг. (соответственно, 283 т). Примечательно, что на этот же период второй половины 1980-х годов приходятся максимальные уловы корюшки в Невской губе и р. Неве (ежегодно в среднем 3 056 т).
Уловы корюшки литовскими промысловиками (данные предоставлены Nature Research Centre и Marin ecology laboratory) в рассматриваемый период времени характеризуются определенной закономерностью: если в 1980-е – начале 1990-х годов ежегодный вылов в среднем не превышал 150 т рыбы (исключая максимальные выловы в 1989 г. – 802 т и 1992 г. – 841 т), то, начиная с 1997 г., показатели вылова достаточно стабильны и не опускаются ниже 300 т (рис. 1).
По данным эстонской рыбохозяйственной науки (официальная статистика предоставлена Estonian Ministry of Environment), объемы вылова корюшки в Нарвском заливе характеризуются относительной стабильностью на протяжении последних лет. Так, в период с 2005 г. по 2008 г. в приграничном с Россией уезде Ида-Вирумаа ежегодные уловы корюшки составили около 16 т. Ранее, в период с 1998 г. по 2004 г., наблюдалось постепенное увеличение промыслового вылова корюшки с максимальными уловами, приходящимися на 2003 г. (34 т) и 2004 г. (33 т).
Данные латышских специалистов рыбного хозяйства (информация предоставлена Institute of Food Safety, Animal Health and Environment «BIOR») свидетельствуют о значительных флуктуациях показателей вылова корюшки в Рижском заливе в период с 1980 г. по 2006 г.: от 648 т в 1980 г. до 10 т в 2002 г. Однако, в последние три года наметился тренд на ежегодное значительное повышение уловов, которые в 2008 и 2009 гг. составили рекордные величины –
1 731 т и 1 941 т, соответственно (рис. 1).
Небезынтересно также привести данные по Пярнусскому заливу Эстонии (который является заливом второго порядка по отношению к Рижскому заливу), где в 2007 и 2008 гг. уловы корюшки составили соответственно 463 т и 628 т (pис. 1). Знаменательно, что оба этих показателя является максимальными значениями вылова после 1984 г. По мнению эстонских специалистов, провал численности корюшки в Пярнусском заливе в 1980-е – первой половине 1990-х годов обусловлен не столько неудовлетворительным состоянием кормовой базы, сколько продолжавшимся в те годы промышленным загрязнением нерестилищ корюшки в реке Пярну в сочетании с неблагоприятными естественными условиями обитания рыбы в этот период (Spilev, Turovski, 2003).
Известно, что на протяжении многих лет промысловые базы (орудия лова корюшки и их количество) в разных странах Балтии остаются достаточно стабильными (с некоторыми оговорками – в отношении Финляндии, где корюшка не представляет существенного коммерческого значения, и интерес к ее добыче у рыбаков неуклонно снижается), а, значит, показатели вылова достаточно точно характеризуют состояние запасов корюшки в разные годы. Разнонаправленные тренды в динамике уловов корюшки в Невской губе, Рижском заливе, Нарвском заливе, вдоль побережья Финляндии и Литвы свидетельствуют о том, что кормовая база у стад корюшки из разных районов моря существенно варьирует.
По-видимому, в условиях дефицита излюбленных кормовых объектов – мизид и гаммарид, рацион питания корюшки стал в большей степени ориентирован не на реликтовую глубоководную фауну, а на молодь рыб и местных беспозвоночных, обитающих на глубинах не более 25-30 м и не затрагиваемых замором, связанным с затоками соленых вод Северного моря (Максимов, 2008).
Предоставляя прекрасные естественные условия для обитания зоопланктона, который сам по себе не является доминирующим в спектре питания взрослой корюшки большую часть года за исключением летних месяцев (Kuhl, 1970; Попов, 1978, 2006а), лагуны и эстуарные зоны Балтики обеспечивают изобильную кормовую базу для питания молоди многих видов рыб, которая, в свою очередь, охотно поедается подрастающей корюшкой. Показано, например, что рыба регулярно встречается в пищевом комке корюшки из восточной части Финского залива, начиная уже с младших размерных классов (9,0-11,9 см) и доминирует в старших размерных классах, причем каннибализм в осенние месяцы может доходить до 100% (Попов, 1978). Схожим образом корюшка из Ботнического залива длиной более 15 см всем другим способам питания предпочитает хищничество (Timola, 1980), а среди рыб, которыми кормится корюшка в эстуарии р. Эльбы, отмечены молодь корюшки, салаки и бычков (Kuhl, 1970).
Для нормального существования многочисленной популяции корюшки с точки зрения ее кормовой обеспеченности, Невская губа предоставляет естественные условия, значительно превосходит потенциал многих эстуарных районов Балтики. Однако, приведенные нами контрастные примеры с динамикой численности популяций корюшки Невской губы и эстуарных районов соседних Прибалтийских стран могут свидетельствовать о том, что несмотря на общее сокращение численности гляциальных реликтов, составляющих известную долю в рационе корюшки Балтики, основные причины ухудшения положения невской корюшки связаны с другими факторами и в первую очередь с глобальным изменением экологической ситуации в Невской губе, – там, где проходит нерест и ранние стадии развития рыбы.
А.Н. Попов (1999, 2004, 2006а) связывает тезис о снижении уровня естественного воспроизводства корюшки с несколькими причинами. Во-первых, «экстремальная гидрометеорологическая обстановка в 1993-1994 гг.» способствовала тому, что «повышенная миграция корюшки в р. Неву приняла устойчивый характер, а нерестилища Южной Лахтинской отмели, где ежегодно воспроизводилось до 40-45% молоди корюшки Финского залива, в настоящее время утратили свое значение». Таким образом, рассуждает автор, запасы корюшки сейчас «пополняются за счет воспроизводства на небольших по площади нерестилищах в Неве и северной части Невской губы». Во-вторых, в Невской губе происходит «сокращение площади нерестовых участков в связи с подводной выемкой грунта и загрязнением». Наконец, в-третьих, «начиная с 1993 г. запасы корюшки пополняются исключительно малоурожайными поколениями».
По нашему мнению, неправильно связывать утрату нерестовых площадей Южной Лахтинской отмели с форс-мажорной гидрометеорологической обстановкой в период 1993-1994 гг. Под «экстремальной» ситуацией А.Н. Попов понимает «образование в конце апреля ярко выраженного температурного градиента на границе зоны транзита невских вод с сильно прогретыми водами южной части Невской губы», что, по мнению автора, «явилось препятствием для нерестовой миграции сюда холодолюбивой корюшки и ее основная масса пошла в Неву».
Инстинкт двигает весенние косяки корюшки в мелководную и распресненную зону залива, где, найдя подходящие нерестовые субстраты, она и мечет икру. Если бы нерестилища Южной Лахтинской отмели продолжали существовать физически, рыба нашла бы к ним путь через неопределенно долгий промежуток времени, в частности, в 1995 г. и в последующие годы, т.е. (по А.Н. Попову) в период «относительно благоприятного развития гидрометеорологических процессов». Если же говорить о причинах физического уничтожения нерестилищ Южной Лахтинской отмели, то это бедствие, прежде всего, связано со вскользь упоминаемым автором публикаций «проведением дноуглубительных работ в Невской губе», которые особенно активно происходят в последнее десятилетие.
Впрочем, гидротехнические работы регулярно проводились в той или иной части акватории Невской губы, начиная еще с Петровских времен, но наиболее масштабные и значительные по воздействию на окружающую среду связаны с возведением дамбы Комплекса защитных сооружений Санкт-Петербурга от наводнений (КЗС), начатым в 1979 г.
Ухудшение экологической ситуации при строительстве дамбы КЗС проявилось, прежде всего, в возникновении значительных площадей малопроточных, застойных зон, что обусловило изменение трофности водоема. Гидробиологические исследования показали значительные колебания трофического статуса экосистемы Невского эстуария за последние 30 лет от олиготрофного до гиперэвтрофного в различных частях эстуария (Голубков и др., 2008б). Эвтрофирование обусловлено повышением концентрации биогенных веществ (фосфора и азота) в малопроточных зонах Невской губы, что способствует быстрому распространению и интенсивному росту колоний цианобактерий, фитопланктона и фитоперифитона. За последние два десятилетия первичная продукция планктона заметно увеличилась практически во всех частях эстуария р. Невы (Голубков и др., 2008а), что, в частности, находит отражение во все более частых и продолжительных периодах «цветения» воды.
Из-за интенсивного эвтрофирования Невской губы происходит вторичное загрязнение прибрежной акватории в результате разложения органического вещества. Основным источником вторичного загрязнения являются в массе размножающиеся нитчатые водоросли рода Cladophora (Никулина, Губелит, 2007). Во время штормов и при колебаниях уровня воды в период смены погоды, нитчатки отрываются от субстрата и скапливаются в мелководных частях губы. В период их разложения отмечается возрастание общего фосфора в воде, снижение прозрачности воды и ухудшение качества придонных местообитаний – гипоксия
(< 3 мгО/л), выделение сероводорода (Березина и др., 2008; Березина, Голубков, 2008; Губелит и др., 2008).
Общее ухудшение экологической ситуации во внутренней акватории Невской губы, начавшееся вскоре после развертывания строительства дамбы КЗС, еще более усугубилось в связи с интенсификацией гидростроительных работ в последние несколько лет (реконструкция Морского порта Санкт-Петербурга, строительство гавани Константиновского дворца в Стрельне, Морского торгового терминала в Ломоносове, «Морского фасада» у западной оконечности Васильевского острова, строительные работы в прибрежье Петергофа, углубление Петровского фарватера и др.).
В результате проведения гидротехнических работ в последние несколько лет территории Южной Лахтинской отмели, также как и значительная часть нерестилищ корюшки Северной Лахтинской отмели, пострадали в большей степени не от выемки грунта (Городилов, 2006; Попов, 2006а), а ровно от противоположного процесса: эти районы, на которых в 1950-1960 гг. добывался строительный песок для нужд города, были определены местами отвалов грунта (или дампинга), изъятого в других районах Невской губы. Складированный таким образом грунт по своей природе преимущественно представляет собой смесь глины и илов четвертичных отложений. В результате действий строителей, не только бывшие песчаные карьеры, но и значительные территории отмелей превратились в заиленные площади, непригодные для икрометания корюшки (Христофоров и др., 2009). Причем в условиях сильного замутнения этих районов, даже уже начавшая развиваться икра рыбы имеет мало шансов на выживание. Икра и молодь рыб наиболее подвержены гибели в результате возрастания концентрации взвешенных частиц грунта из-за невозможности быстро покинуть замутненные участки (Лаврентьева, Суслопарова, 2006).
Известно, что более 70% загрязняющих веществ, поступающих со стоком реки Невы, а также через станции аэрации или с ливневыми стоками, оседает в пределах Невского эстуария (Рыбалко, Федорова, 2008). Именно по этой причине начало наиболее масштабных гидротехнических работ в Невской губе в 2005 г. совпадает со временем резкого ухудшения экологической ситуации по индексу загрязненности придонных вод Невского эстуария тяжелыми металлами и нефтепродуктами, которые многие десятилетия аккумулировались в донных осадках и при строительстве были переведены во взвешенное состояние (Федорова и др., 2008). На спутниковых снимках Восточной части Финского залива последних лет (Булаев, Чичкова, 2008; Сухачева и др., 2008) отчетливо видно, что зоны наиболее интенсивной замутненности воды в весеннее и летнее время расположены на участках производства гидротехнических работ (в т.ч. в районах Южной и Северной Лахты) и примыкающих к ним и к дамбе КЗС акваториях, однако повышенное осадконакопление распространяется со шлейфами мути далеко за пределы Невской губы (Рыбалко и др., 2009; Чичкова, Минаков, 2010) (рис. 2). Примечательно, что по сравнению с 2005 г., когда в акватории восточной части Финского залива преобладали незначительные загрязнения, близкие к фоновым показателям, в период 2007-2009 гг. преимущественно отмечались умеренные, сильные (зона повышенной мутности распространяется более чем 50% площади анализируемой акватории) и максимальные (зона повышенной мутности закрывает 100% площади анализируемой акватории) стадии загрязнения (табл. 1).
Таблица 1. Распределение зон повышенной мутности в восточной части Финского залива по стадиям загрязнения (по Чичкова, Минаков, 2010).
Table 1. The distribution of zones of water turbidity in the Eastern part of the Finnish Gulf according to the stages of pollution (follow to Chichkova, Minakov, 2010).
| Стадии загрязнения | 2005 г. | 2007 г. | 2008 г. | >2009 г. | Всего с 2005 по 2009 гг. |
| Незначительное | 18 | 2 | 4 | 6 | 30 |
| Умеренное | 10 | 6 | 14 | 12 | 42 |
| Сильное | 0 | 5 | 9 | 8 | 22 |
| Максимальное | 0 | 7 | 3 | 1 | 11 |
Антропогенное влияние и сопряженное с ним изменение трофического статуса водоема обусловливают не только падение запасов холоднолюбивых рыб, но и отражается на ухудшении физиологического состояния, восприимчивости рыб к болезням. Ввиду способности рыб к аккумуляции тяжелых металлов, концентрация их в мышцах и, главным образом, во внутренних органах отражает реальное состояние водного объекта, что позволяет использовать рыб для индикации загрязнения водных экосистем. Проведенное в 2009 г. в лаборатории экологической токсикологии ГосНИОРХ Е.С. Светашовой исследование показало, что концентрации свинца в мышцах корюшки, выловленной на выходе из Невской губы (р-н г. Сестрорецк) в 2,7 раза, а меди – в 1,7 раза выше, чем в мышцах корюшки из Нарвского залива. Значительное накопление тяжелых металлов наблюдалось в почках анализируемых рыб, причем, в почках корюшки из р-на Сестрорецка концентрация всех токсикантов была выше, чем в почках рыб из Нарвского залива, а именно: цинка – в 1,4 раза, кадмия – в 2,1 раза, свинца – в 2,4 раза, меди – в 1,2 раза.
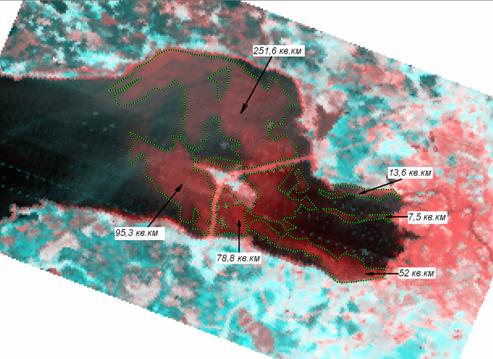
Рис. 2. Площади повышенной мутности в зоне влияния комплекса защитных сооружений,
30 мая 2009 г. (по Чичкова, Минаков, 2010).
Fig. 2. Squares of water turbidity in the region of dam influence. May, 30, 2009 (follow to Chichkova, Minakov, 2010).
Известно, что в условиях повышения трофности водоема, как на гидробионтах, так и в среде их обитания возрастает встречаемость патогенных микроорганизмов. Исследования по оценке встречаемости возбудителя пищевого листериоза (бактериальной инфекции с высокой степенью летальности для человека) показали, что в водах прибрежной зоны Финского залива встречаемость сапрофитных листерий вида Listeria monocytogenes составляет 14,3%, а патогенных 21,4%. На свежевыловленной корюшке из Финского залива показатель встречаемости сапрофитных листерий составил 30,8%, а патогенных – 38,5%. Примечательно, что наибольшие концентрации листерий в пробах были обнаружены в непосредственной близости от выхода из Невской губы. Так, 100% проб рыбы, выловленные в устьевой части р. Сестры, содержали L. monocytogenes, при этом все пробы воды из этого же района содержали патогенный вид листерий
(Байдова и др., 2009).
Вернемся к проблеме малоурожайных поколений, за счет которых, по свидетельству А.Н. Попова (1999, 2004, 2006а), стали пополняться запасы корюшки, начиная с 1993 г. Автор публикаций связывает это явление с сокращением численности в Финском заливе энергетически ценных ракообразных, а также с прекращением нереста корюшки в южной части Невской губы в связи с неблагоприятными гидрометеорологическими условиями. Однако, по нашему мнению, сокращение численности популяции Невской корюшки происходило, в том числе, не без прямого и косвенного влияния дамбы, начиная с первых лет ее строительства. Так, построенная раньше южного участка северная ветка КЗС явилась труднопреодолимым препятствием для идущих на нерест косяков корюшки. Возможно, сильные выносные течения из водопропускников (в отсутствии сильных нагонных ветров западных румбов скорости течения достигали нескольких метров в секунду) служили сигналом к запрету дальнейшей нерестовой миграции не самого реофильного вида рыб, либо корюшка просто не находила оставленные в дамбе проходы (к сведению: общая ширина всех водопропускных (6 шт.) и судопропускных (2 шт.) сооружений построенной дамбы – 1,8 км, что составляет около 8% от общей протяженности водной части дамбы – 22,2 км.). Не только корюшка, но и рыбы с более ярко выраженными навигационными способностями не могли преодолеть оставленные узкие проходы в дамбе: например, известны случаи, когда местные жители отлавливали по ночам массово переползающих дамбу угрей во время их катадромной миграции, и понадобилось вмешательство милиции, чтобы пресечь эту форму браконьерства.
До строительства дамбы КЗС основной объем стока Невских вод проходил вдоль северного берега Невской губы, в створе между Горской и Котлином. На противоположном берегу залива преобладающее направление движения вод имело обратный вектор, и было направлено к городу. После перекрытия северного участка дамбы в 1984 г. глобальная циркуляция вод в Невской губе была кардинально нарушена, сток из Невы был по существу перенаправлен в открытый створ
Котлин – Бронка и это могло в значительной степени повлиять на быстрый выход из строя нерестилищ Южной Лахтинской отмели. Зона перманентного застоя воды, образовавшаяся в этом районе, способствовала быстрому осадконакоплению и заилению нерестилищ. Слабая выживаемость икринок и личинок корюшки на ранних этапах жизненного цикла могла быть связана со стремительным весенним прогревом застойных вод Южной Лахты, а также с качественным и количественным нарушением устойчивости сообщества зоопланктона – основы питания корюшки на личиночной стадии. Результаты гидробиологических исследований, проведенных на прилегающих к южному побережью Невской губы акватории в период с 1983 по 1993 гг., показали, что величина индекса видового разнообразия зоопланктона в те годы варьировала от 2,57 до 3,53 и имела тенденцию к снижению (Иванова, Телеш, 1996). Те же индексы, рассчитанные для подводного отвала на Южной Лахтинской отмели и на участках в районах гавани пос. Стрельна в 2002 г. были значительно ниже и не превышали значения 1,07. При этом из сообщества зоопланктона в первую очередь элиминировались мелкие формы, жизненно важные на начальных этапах питания корюшки (Суслопарова и др., 2006).
Кроме того, известно, что до строительства дамбы личинки корюшки, которая нерестилась в реке Неве и у северного побережья Невской губы, беспрепятственно выносились в залив с транзитными невскими водами. По данным А.Н. Попова (2006а), наибольшая концентрация этих личинок в поверхностном слое воды обнаруживалась в июне в районе действия турбулентных течений у Сестрорецка-Зеленогорска, где продолжалось их нормальное развитие с переходом в мальковую стадию. После завершения основного строительства обеих веток дамбы, водообмен между акваториями по обе стороны от КЗС изменился, и это не могло не повлиять на привычный режим пассивной миграции личинок корюшки в открытую часть залива.
Считается, что водообмен Невской губы после завершения основного строительства дамбы остался достаточно высоким, период пребывания воды составляет в среднем 5,5 суток, а в центральной транзитной зоне – вдвое меньше (Балушкина, 2008). Однако для районов Невской губы, находящихся на периферии от транзитных участков, этот показатель заметно ниже. Косвенным образом об этом могут свидетельствовать интегральные показатели качества вод эстуария р. Невы в 1994-2004 гг.: если для вод транзитных зон с наибольшими скоростями проточности качество вод было оценено как «умеренно загрязненные», то для вод большей части Невской губы оценивалось как «загрязненные» (Алимов, Голубков, 2008). В этой связи можно предполагать, что для личинок корюшки, которые дрейфуют с речным потоком с нерестилищ из р. Невы, время пребывания в Невской губе изменилось незначительно. Однако, по аналогии с Южной Лахтой, значительное количество личинок с северного побережья Невской губы на пути к дамбе теперь попадает в застойные зарослевые зоны (наибольшее количество районов с подобными условиями наблюдается на расстоянии 3-5 км от дамбы) и не добирается до водо- и судопропускников. В норме личинки корюшки никогда не встречаются в подводных зарослях, они принадлежат к альтернативному – пелагическому биоценозу личинок Невской губы, спектр питания которых ориентирован на характерные для данного биотопа зоопланктонные организмы – ветвистоусых, веслоносых рачков, коловраток (Огородникова, Суслопарова, 1986). Таким образом, большая часть попавших в зарослевый биоценоз личинок корюшки страдает от недостатка корма и подвержена действию асфиксии: на примере обширного участка зарослей макрофитов северной ветки дамбы показано, что при высокой биомассе зообентоса и молоди рыб биомасса зоопланктона здесь была близка к минимальной (Лаврентьева и др., 2006). Остается лишь с сожалением констатировать, что после возведения дамбы КЗС участков с зарослями полупогруженной растительности стало больше (Жакова, 2008; Ковальчук, 2008), причем экспансия зарослей продолжается, особенно в прилегающих к дамбе вдольбереговых районах слабопроточных прибрежных вод.
Негативное влияние на зоопланктон и личинок корюшки, дрейфующих с нерестилищ Северной Лахты и из Невы, усугубляется в связи со значительным повышением мутности воды в результате гидростроительства: при фоновых значениях средней мутности вод Невской губы 7 мг/л, мутность в местах дноуглубления и дампинга превышает 100 мг/л. Известно, что резкое возрастание концентрации взвешенных частиц грунта в воде приводит к гибели водных животных-фильтраторов – как планктонных, так и бентосных. Планктон погибает от потери плавучести и асфиксии. Часть живых организмов оседает на дно и гибнет при налипании на них частиц грунта (Лаврентьева, Суслопарова, 2006).
Показано, что в зоне дноуглубления на участках с максимальными концентрациями грунтовой взвеси обилие (численность, биомасса, продукция) зоопланктона в целом снижалось в 3-5 раз по сравнению с фоновыми показателями. В зоне дампинга при взятии проб сразу после сброса и непосредственно на его участке отмечалась практически 100%-ная гибель зоопланктона (Лаврентьева, Суслопарова, 2006). Схожая тенденция наблюдалась и по фитопланктону: в 2006-2007 гг. водоросли планктона были представлены 6-10 видами, при этом биомасса была в 10-15 раз ниже обычно наблюдаемой по Невскому эстуарию (Никулина, 2008). Поскольку дамба КЗС ограничивает свободный водообмен между Невской губой и открытым заливом, негативное влияние от взмученной воды на биоту губы продлевается как во времени, так и в пространстве, воздействуя практически на всю ее внутреннюю акваторию, что хорошо видно по спутниковым фотографиям последних лет. Кроме того показано, что пролонгирование негативного воздействия замутнения возникает также из-за мелководности Невской губы: в начале нового вегетационного сезона наблюдается вторичное замутнение акватории даже в тех ее районах, которые находятся в зонах транзита невских вод и активного водообмена (Суслопарова и др., 2008).
Исследованиями последних лет показано, что гидротехнические работы являются основным фактором, лимитирующим биопродуктивность фито-, зоопланктона, макрозообентоса и ихтоифауны на отдельных участках Невской губы (Суслопарова и др., 2009). И без того заметно снизившаяся в последние годы концентрация личинок корюшки на ее исконных нерестилищах только за два года наблюдений (2006-2007) упала в районе Северной Лахты в 9 раз (с 2,14 экз./м3 до 0,23 экз./м3), в районе Южной Лахты в 30 раз (с 7,64 экз./м3 до 0,25 экз./м3) и районе западнее Васильевского острова в 20 раз (с 6,11 экз./м3 до 0,30 экз./м3) (Суслопарова и др., 2008).
Однако, даже если бы в пределах Невской губы мутность оставалась на фоновом уровне, в водопропускниках дамбы личинки корюшки имеют повышенный риск быть употребленными в пищу всеми видами рыб, обитающими в Невской губе. Водо- и судопропускные сооружения по сути представляют собой короткие неширокие реки, в которые канализируется весь сток из Невской губы вместе с пассивно дрейфующим в нем фито- и зоопланктоном. По аналогии с реками, концентрация активно питающейся рыбы на участках с выраженным течением (непосредственно в водопропускниках дамбы, а также на входе и выходе из них) многократно превышает усредненную концентрацию рыб в водоеме (в случае с Невской губой – в створе между коренным берегом и о. Котлин), поэтому шансы личинок корюшки благополучно миновать дамбу заметно снижаются. Таким образом, в связи со строительством дамбы уязвимость корюшки на самых ранних этапах жизненного цикла (икринки и личинки) увеличилась многократно.
Примечательно, что ни в одной из своих статей А.Н. Попов не признает негативного влияния дамбы на популяцию невской корюшки. Его аргументация строится исключительно на данных анализа уловов нерестовой корюшки на разных промысловых участках до и после постройки дамбы. Вывод автора статей однозначен: «относительные величины уловов после перекрытия северного участка (декабрь 1984 г.) вплоть до 1993 г. в целом сохраняются, что свидетельствует о беспрепятственной миграции корюшки через водо- и судопропускные отверстия защитных сооружений в Невскую губу и р. Неву» (Попов, 2006а).
Однако, если исходить из приводимых автором статьи цифр (Попов, 2006а), начиная с 1993 г., картина существенно меняется: на Сестрорецком участке промысла, т.е. со стороны моря, корюшки добывается все больше относительно уловов во внутренней акватории Невской губы (на участке Лахта – Лисий Нос по северному берегу). Данные по уловам в статье А.Н. Попова заканчиваются 2002 г., однако смещение в распределении уловов подтверждается цифрами последних лет, при этом отмеченная выше тенденция становится еще более очевидной (табл. 2).
Сами по себе рассуждения, построенные на данных о величине относительных уловов как таковых, вызывают большой скепсис, поскольку со времени перехода к новым экономическим отношениям в конце 1980-ых годов, рыбохозяйсвтенная статистика практически перестала отражать реально существующее положение дел в рыболовстве, причем регулирование промысла и контроль за ним со стороны государства полностью не восстановлены по настоящее время.
Неучтенный вылов (под ним понимается нелегальный промысел, а также вылов рыбы сверх разрешенных лимитов законно хозяйствующими на водоемах организациями) является одной из важнейших причин угасания популяции невской корюшки. Наибольший размах это явление получило в последнее десятилетие в связи с доступностью и крайне низкой ценой на сетные орудия лова. В сезон нерестового хода корюшки вся акватория Невской губы буквально опутана браконьерскими сетями, причем наибольшая концентрация незаконно выставленных орудий лова наблюдается в районе КЗС. Сотни километров сетей, мережи, заколы выставляются там, где рыбу проще всего взять, т.е. вдоль тела дамбы. Максимальная концентрация браконьерских снастей образуется в районах, примыкающих к судо- и водопропускным отверстиям. Шансы миновать частокол сетных преград у корюшки крайне малы. Отсутствие надлежащего контроля на водоеме в период проведения корюшковой путины объясняется как общеизвестными недостатками в организации рыбоохранных мероприятий в нашей стране, так и чрезвычайно высокой коррупционной емкостью корюшкового бизнеса, поскольку цена на корюшку уже давно превысила цену на судака, сига или лосося… Таким образом, губительное воздействие КЗС на популяцию корюшки опосредованно реализуется и через незаконную рыбодобычу, которая чрезвычайно возросла в районе дамбы.
Таблица 2. Среднегодовые уловы нерестовой корюшки по участкам промысла (данные предоставлены Севзапрыбводом).
Table 2. Annual catches of adult smelt in different sites of fishery (data provided by Sevzaprybvod).
| Годы | Показатели | Сестрорецк | Стрельна | Лахта - Лисий Нос | р. Нева | Всего |
| 1980-1985 | Уловы, т | 403 | 802 | 308 | 191 | 1704 |
| % | 24 | 47 | 18 | 11 | 100 | |
| 1986-1992 | Уловы, т | 795 | 1000 | 421 | 301 | 2517 |
| % | 31 | 40 | 17 | 12 | 100 | |
| 1993-1997 | Уловы, т | 339 | 69 | 110 | 187 | 705 |
| % | 48 | 10 | 16 | 26 | 100 | |
| 1998-2002 | Уловы, т | 172 | 30 | 62 | 124 | 388 |
| % | 44 | 8 | 16 | 32 | 100 | |
| 2003-2004 | Уловы, т | 36 | 8 | 7 | 21 | 72 |
| % | 50 | 11 | 10 | 29 | 100 | |
| 2005 | Уловы, т | 6,1 | 2,8 | 1,5 | 7,9 | 18,3 |
| % | 34 | 15 | 8 | 43 | 100 | |
| 2006 | Уловы, т | 44,7 | 5,4 | 6,9 | 23,7 | 80,7 |
| % | 55 | 7 | 9 | 29 | 100 |
|
| 2007 | Уловы, т | 34,8 | 30,0 | 4,0 | 25,9 | 94,7 |
| % | 37 | 32 | 4 | 27 | 100 | |
| 2008 | Уловы, т | 31,5 | 14,0 | 1,6 | 14,6 | 61,7 |
| % | 51 | 23 | 2 | 24 | 100 |
Очевидно, что пока промысловое изъятие в любом его виде не превышает расчетную допустимую величину улова, использование тех или иных орудий лова при промысле корюшки не имеет решающего значения. Поэтому тезис о том, что «использование на промысле корюшки в основном пассивных орудий лова типа ловушек: корюшковых заколов, ставных неводов и мереж – не оказывает отрицательного воздействия на снижение уровня ее естественного воспроизводства» (Попов, 2006а) может быть принят лишь при условии недопущения перелова.
Известно, что с ухудшением качества воды, условий нагула, питания, и размножения гидробионтов как в структуре зоопланктона, так и в структуре рыбного населения происходит замещение крупных и долгоживущих форм на формы мелкие и раносозревающие. Этот процесс наглядно демонстрируется изменением состава популяции невской корюшки в последнее десятилетие: из-за сокращения возрастного ряда процент особей старших возрастов в стаде уменьшается, рыба раньше становится половозрелой и тем самым все больше напоминает корюшку из Ладоги или из Выборгского залива. В целом процесс измельчания корюшки сопровождается увеличением ее продукции и биомассы, но как следствие типичная невская корюшка с относительно длинным жизненным циклом заменяется на «сорную» мелкую рыбу с высоким уровнем воспроизводства и высоким приростом продукции.
В связи с общим снижением численности невской корюшки, а также с происходящим в настоящее время заметным ее измельчением наблюдается очевидное уменьшение пресса любительского рыболовства на корюшку Финского залива. В отличие от корюшковой рыбалки 1970-1990-ых годов, имевшей статус социального явления для многомиллионного Санкт-Петербурга и жителей Ленинградской области, нынешний лов этой рыбы любительскими снастями стал малоинтересен и попросту нерентабелен.
Таким образом, заключение А.Н. Попова о том, что «на фоне снижения численности корюшки отмечается небывалый рост любительского подледного лова в Финском заливе, отрицательное воздействие которого на запасы корюшки в настоящее время не поддается учету», мало соответствует действительности.
К сожалению, ошибочность вывода автора публикаций о резко негативном влиянии любительского зимнего рыболовства не так безобидна, как могла бы показаться на первый взгляд. Дело в том, что экспертная оценка величины вылавливаемой любителями рыбы используется при расчете общего допустимого улова (ОДУ) – показателя, на основании оценок которого рассчитываются перспективные ежегодные квоты на вылов биологических ресурсов в водоеме.
Расчет ОДУ проводится по модели ВПА (виртуально-популяционного анализа), в которую в качестве исходных данных независимо друг от друга должны закладываться величины промыслового и любительского уловов. При ошибочном подходе, применяемом в настоящее время, значение величины любительских уловов суммируются к ОДУ, полученному только по данным промысловой статистики.
Кроме того, по экспертным оценкам автора производимых расчетов, ежегодные уловы любителей в последние годы составляют минимум четверть от всей изымаемой корюшки. Со справедливостью этой оценки трудно согласиться, поскольку при преобладающих в последние годы мягких зимах условия для подледной рыбалки на Финском заливе складывались далеко не каждый год.
Таким образом, результирующее значение ОДУ заведомо оказывается завышенным. А это означает, что промыслом систематически изымается рыбы больше, чем это возможно для нормального существования популяции корюшки. Промышленный перелов приводит к биологическому перелову – подрыву запаса, при котором восстановить численность популяции до исходной величины без принятия специальных мер по ограничению вылова затруднительно.
Косвенным образом на биологический перелов невской корюшки указывает сокращение возрастного ряда нерестового стада рыбы, наблюдающееся в последние годы.
Резюмируя вышесказанное, можно заключить, что малоурожайность поколений корюшки объясняется многими причинами, среди которых необходимо выделить следующие основные:
- Ухудшение кормовой базы нагуливающейся в заливе рыбы.
- Влияние дамбы КЗС, прямо и опосредованно воздействующей как на количество заходящих в губу на нерест рыб, так и на выживаемость и выход из губы личинок корюшки.
- Масштабные гидротехнические работы в Невской губе, которые в связи с сопутствующим им загрязнением являются причиной систематического вывода из оборота нерестилищ корюшки и ухудшения выживаемости оплодотворенной икры и личинок корюшки.
- Возрастающий пресс браконьерского лова.
- Некорректный расчет величины ОДУ, приводящий к выдаче завышенных квот на рыбодобычу, что неизбежно приводит к последующему перелову нерестового стада.
Очевидно, из всех негативных факторов определить лимитирующий невозможно: каждый из них, будучи весьма важным сам по себе, умножает действие других при их комплексном воздействии. Однако, если тенденция снижения количества основных объектов питания корюшки в заливе определяется глобальными климатическими процессами, то остальные факторы относятся к числу антропогенных, а значит, по крайней мере, поддающихся корректировке, если не полному устранению.
Поскольку уже в настоящее время популяция невской корюшки вынуждена обитать в условиях, значительно отличающихся от экологической ситуации на момент до 1979 г., после окончания достройки дамбы КЗС должны быть соблюдены условия, максимально обеспечивающие беспрепятственную нерестовую миграцию производителей и свободную пассивную миграцию личинок корюшки. Любой вид промыслового лова должен быть вынесен за пределы примыкающей к дамбе акватории на оговоренное расстояние. Основной объем гидротехнических работ на внутренней акватории Невской губы следует проводить в сроки, не совпадающие с временем нереста и развития личинок корюшки (весной и в начале лета), при этом работа техники должна быть организована таким образом, чтобы при минимальных повышениях мутности воды на акватории залива (установка улавливающих муть тканевых экранов отчасти позволяет этого добиться) обеспечивалась сохранность традиционных нерестилищ корюшки в Невской губе.
С учетом переформирования рельефа дна и береговой линии в связи с проведением интенсивных работ на акватории Невской губы, следует пересмотреть результаты предшествующих экспериментальных и теоретических исследований по оценке и прогнозу прохождения транзитных вод в Невской губе, поскольку в районах с измененной батиметрией поля течений могут существенно изменяться (Кондратьев и др., 2009). Так, несмотря на полученное в свое время положительное заключение экологической экспертизы строительства КЗС, которое было основано на многочисленных теоретических выкладках и модельных построениях, возникшие на практике обширные зоны застойных вод (особенно вдоль северного берега) красноречиво свидетельствуют об ухудшении экологической ситуации в Невской губе. Очевидно, что по крайней мере для частичного решения этой проблемы потребуется увеличить глубину створов двух первых водопропускников (В5 и В6) по северной ветке дамбы.
В качестве рекомендаций по восстановлению численности невской корюшки следует особо отметить необходимость проведения рекультивации нерестилищ на территориях Южной и Северной Лахты. Для этого требуется подсыпка песка на площадях, которые ранее служили нерестилищами и теперь выведенные из оборота из-за дампинга моренного грунта. Опыт Северной Лахты показал, что больших материальных ресурсов для этого не потребуется: по данным, поступившим от профессиональных рыбаков, на небольших участках Северной Лахты, куда в 2007 г. поверх отвалов глинистых грунтов был выгружен песок, корюшка нерестилась уже весной 2008 г. Очевидно, что основная техническая задача при проведении подобного рода мероприятий – удержание от смывания течением песка на месте его выгрузки, чего, по-видимому, можно достичь установкой в грунт шпунтов.
Территориям Северной и Южной Лахты, как районам наиболее важным для воспроизводства невской корюшки, следовало бы придать статус рыбоохранных заказников. Ранее в рамках Генплана Санкт-Петербурга 2005 г. уже предлагался проект создания ООПТ «Ихтиологический заказник «Питерская корюшка»», площади которого (всего порядка 250 га) частично бы занимали территории Северной и Южной Лахты. Очевидно, что в случае реализации этого крайне важного для поддержания невской корюшки проекта площади охраняемых территорий должны быть скорректированы в сторону их увеличения.
Особый режим рыбохозяйственного использования также должен быть определен для Канонерской отмели – еще одного важного района нереста корюшки и других видов рыб. Несмотря на близость к Санкт-Петербургу, дноуглубительные работы на Канонерской отмели не проводились с позапрошлого века. Там до сих пор обитают многие виды гидробионтов, не встречающиеся в других местах Невской губы и среди них такие индикаторы чистоты, как раки и пескари.
Отдельное внимание при восстановлении запасов корюшки должно быть уделено изучению потенциальной емкости нерестилищ этой рыбы, расположенных за пределами Невской губы. Известно, что в восполнении запасов полупроходной корюшки восточной части Финского залива определенную роль может играть Сестрорецкая отмель, реки (например, Нарова, Луга, Систа, Сестра), и эстуарные части некоторых из них, однако данные об этом весьма скудны, инвентаризационные исследования площадей нерестилищ не проводились.
Усугубившаяся в последние годы тенденция снижения запасов корюшки в заливе заставляет специалистов задуматься о возможности искусственного воспроизводства этой рыбы. В настоящее время разрабатываются проекты по подращиванию корюшки в бассейнах вплоть до товарной рыбы, однако более перспективными кажутся предложения по инкубации икры и подращиванию корюшки до стадии личинки (Городилов, 2006), которую затем планируется выпускать в залив западнее дамбы КЗС. Известно, что в предстоящие годы в районе Сестрорецкой отмели, а также в северной части о. Котлин намечены новые грандиозные гидротехнические работы (отсыпка не существующих пока островов на участке Тарховской бухты и вдольбереговой зоны западнее пос. Александровская, строительство здесь яхтинга с постоянным изъятием акватории не менее 200 га; значительное увеличение площади о. Котлин за счет намыва территории в северной его части для размещения учебной базы ВМФ; в течение ближайших шести лет добыча песка из подводных карьеров в акватории напротив г. Сестрорецка), которые будут сопровождаться неизбежным ухудшением экологической ситуации в зоне турбулентных вод в этом районе, а, значит, снизит выживаемость концентрирующихся здесь личинок невской корюшки. Однако, принимая во внимание сложившиеся на сегодняшний день бедственные экологические условия в Невской губе, без мер по поддержанию популяции корюшки за счет ее искусственного воспроизводства, по крайней мере, до стадии подрощенной личинки, обойтись будет трудно.
В заключении следует отдельно остановиться на проблеме определения общего допустимого улова корюшки. Расчисление величины ОДУ невской корюшки должно проводиться с учетом и трезвой оценкой всех множественных факторов, влияющих на численность ее популяции. Принимать во внимание необходимо даже те, казалось бы, второстепенные обстоятельства, с которыми корюшки имеет потенциальные шансы столкнуться на разных этапах своего жизненного цикла, вплоть до учета потерь дрейфующих по Неве икринок и личинок корюшки на главном водозаборе Санкт-Петербурга или, например, на насосных установках плавучего фонтана на Стрелке Васильевского острова (если он вновь будет функционировать). Биологический смысл происходящих с популяцией корюшки процессов не должен теряться в формулах сложной, но не вполне адекватной математической модели, применяемой сейчас при расчете ОДУ. При этом при получении статистических оценок предпочтение должно отдаваться реальным (а не виртуальным) первичным данным, основанных на методах прямого учета.
Специалисты по прогнозированию запасов рыб полагают, что уже в ближайшее время понижающаяся сейчас ветвь долгопериодного цикла продуктивности Балтийского моря изменит знак на противоположный, и запасы корюшки вновь начнут возрастать (Кудерский и др., 2008). К сожалению, в условиях радикального ухудшившейся экологической ситуации в Невской губе и отсутствия обоснованных надежд на скорое улучшение этого положения объективный прогноз на перспективы невской корюшки менее оптимистичен. Очевидно, в ближайшие годы последствия перелова заметно оскудевшего стада невской корюшки, утратившей нормальные условия для нереста и ранних этапов развития в Невской губе, приведут к дальнейшему значительному снижению численности ее популяции. В случае реализации этого печального сценария, потребность петербуржцев в корюшке будет все больше восполняться за счет дешевого эрзаца в виде привозной замороженной Азиатской зубатой корюшки, Osmerus mordax dentex, которая даже по своему систематическому статусу находится в весьма отдаленном родстве с корюшкой Европейской (Сендек и др., 2005), а для спасения некогда главной промысловой рыбы Невской губы придется принимать беспрецедентные меры, вплоть до объявления полного запрета на ее вылов. Однако, даже при менее драматичном развитии событий, без решения системной проблемы грамотного расчета ОДУ для невской корюшки прочие меры по восстановлению численности самой популярной рыбы Санкт-Петербурга заведомо обречены на неудачу.
Благодарности
Авторы выражают искреннюю признательность коллегам из Прибалтийских стран, предоставивших сведения по промысловой статистике корюшки: Маркку Каукоранте (Markku Kaukoranta, Финляндия), Витаутасу Кесминасу и Римантасу Репечке (Vytautas Kesminas, Rimantas Repecka, Литва), Маркусу Виетемаа (Markus Vetemaa, Эстония), Янису Бирзаксу (Janis Birzaks, Латвия). Отдельная благодарность Е.Ф. Чичковой за разрешение использовать в статье авторские материалы по спутниковому мониторингу за восточной частью Финского залива.
СПИСОК ЛИТЕРАТУРЫ
Алимов А.Ф., Голубков С.М. Роль естественных и антропогенных факторов в современной динамике эстуария р. Невы и стратегия управления его биологическими ресурсами. Сб. Экосистема эстуария реки Невы: биологическое разнообразие и экологические проблемы / Под ред. А.Ф. Алимова, С.М. Голубкова. М.: Изд-во «Товарищество научных изданий КМК», 2008. С. 427-437.
Байдова Т.В., Вишнякова Л.А., Приймак Л.Я., Репина О.И. Оценка встречаемости Listeria monocytogenes в корюшке европейской и воде Финского залива. Сб. День Балтийского моря: тезисы докл. X Международного экологического форума (СПб, 17-19 марта 2009 г.). СПб., 2009. С. 150-151.
Балушкина Е.В. Оценка состояния экосистемы и качества вод эстуария реки Невы по показателям зообентоса. Сб. Экосистема эстуария реки Невы: биологическое разнообразие и экологические проблемы / Под ред. А.Ф. Алимова, С.М. Голубкова. М.: Изд-во «Товарищество научных изданий КМК», 2008. С. 411-426.
Балушкина Е.В., Голубков С.М., Голубков М.С., Максимов А.А. Роль антропогенных факторов в динамике сообществ зообентоса. Сб. Экосистема эстуария реки Невы: биологическое разнообразие и экологические проблемы / Под ред.
А.Ф. Алимова, С.М. Голубкова. М.: Изд-во «Товарищество научных изданий КМК», 2008. С. 356-369.
Березина Н.А., Голубков С.М. Динамика зообентоса прибрежной зоны в условиях эвтрофирования. Сб. Экосистема эстуария реки Невы: биологическое разнообразие и экологические проблемы / Под ред. А.Ф. Алимова, С.М. Голубкова. М.: Изд-во «Товарищество научных изданий КМК», 2008. С. 370-383.
Березина Н.А., Голубков С.М., Губелит Ю.И., Балушкина Е.В., Ципленкина И.Г. Состояние литоральных биоценозов восточной части Финского залива (1998-2007 гг.). Сб. День Балтийского моря: тезисы докл. IX Международного экологического форума (СПб, 11-13 марта 2008 г.). СПб.: Изд-во «Диалог», 2008. С. 219-220.
Булаев О.А., Чичкова Е.Ф. Спутниковый мониторинг восточной части Финского залива в 2007 г. Сб. День Балтийского моря: тезисы докл. IX Международного экологического форума (СПб, 11-13 марта 2008 г.). СПб.: Изд-во «Диалог», 2008. С. 224.
Галкина Н.П., Широков Л.В. О распределении и численности личинок рыб в восточной части Финского залива // Изв. ГосНИОРХ. 1978. Т. 129. C. 43-52.
Голубков М.С., Голубков С.М., Умнова Л.П. Первичная продукция и проблемы эвторфирования эстуария р. Невы. Сб. Экосистема эстуария реки Невы: биологическое разнообразие и экологические проблемы / Под ред. А.Ф. Алимова, С.М. Голубкова. М.: Изд-во «Товарищество научных изданий КМК», 2008а. С. 313-337.
Голубков М.С., Голубков С.М., Умнова Л.П. Факторы, влияющие на колебания трофического статуса эстуария р. Невы и Выборгского залива в течение последних десятилетий. Сб. День Балтийского моря: тезисы докл. IX Международного экологического форума (СПб, 11-13 марта 2008 г.). СПб.: Изд-во «Диалог», 2008б. С. 63-64.
Городилов Ю.Н. О состоянии популяции корюшки европейской Osmerus eperlanus eperlanus (L.) в Невской губе Финского залива. Сб. Экология г. Петергофа – Наукограда Российской Федерации и сопредельных территорий: тезисы Международной научно-методической конференции. СПб., 2006. С. 22-23.
Губелит Ю.И., Голубков С.М., Березина Н.А., Никулина В.Н. Проблемы эвтрофирования прибрежных вод эстуария реки Невы. Сб. Экосистема эстуария реки Невы: биологическое разнообразие и экологические проблемы / Под ред. А.Ф. Алимова, С.М. Голубкова. М.: Изд-во «Товарищество научных изданий КМК», 2008. С. 338-345.
Жакова Л.В. Макрофиты: высшие водные растения и макроводоросли. Сб. Экосистема эстуария реки Невы: биологическое разнообразие и экологические проблемы / Под ред. А.Ф. Алимова, С.М. Голубкова. М.: Изд-во «Товарищество научных изданий КМК», 2008. С. 105-125.
Иванова М.Б., Телеш И.В. Оценка экологического состояния Невской губы и водотоков С.-Петербурга по зоопланктону. Сб. Экологическое состояние водоемов и водотоков бассейна реки Невы. СПб., 1996. С. 36-52.
Ковальчук Н.А. Макроводоросли как индикатор процесса эвтрофирования прибрежных вод российской части Финского залива. Сб. День Балтийского моря: тезисы докл. IX Международного экологического форума (СПб, 11-13 марта 2008 г.). СПб.: Изд-во «Диалог», 2008. С. 253.
Кожевников Г.П. Биология и экология невской корюшки Osmerus eperlanus (L.): диссерт. на соиск. ученой степени канд. биол. наук. Л., 1949. 186 с.
Кондратьев С.А., Рябченко В.А., Дворников А.Ю. и др. Изменение течений и траекторий распространения шлейфов сточных вод на акватории Невской губы в результате переформирования береговой линии и дна. Сб. День Балтийского моря: тезисы докл. X Международного экологического форума (СПб, 17-19 марта 2009 г.). СПб.: Изд-во ООО «Макси-Принт», 2009. С. 54-55.
Кудерский Л.А. Что происходит с рыбными запасами в восточной части Финского залива: есть ли связь со строительством защитных сооружений г. Санкт-Петербурга от наводнений // Жизнь и безопасность. 2005. №1-2. С. 412-417.
Кудерский Л.А., Шурухин А.С., Попов А.Н., Богданов Д.В., Яковлев А.С. Рыбное население эстуария реки Невы. Сб. Экосистема эстуария реки Невы: биологическое разнообразие и экологические проблемы / Под ред. А.Ф. Алимова, С.М. Голубкова. М.: Изд-во «Товарищество научных изданий КМК», 2008. С. 223-240.
Лаврентьева Г.М., Суслопарова О.Н. Итоги рыбохозяйственных мониторингов, проводимых в восточной части Финского залива с целью оценки воздействия гидротехнических работ на гидробионтов // Сб. науч. тр. ГосНИОРХ. 2006. Вып. 331. Т. 1. С. 5-11.
Лаврентьева Г.М., Суслопарова О.Н., Аршаница Н.М. и др. Характеристика современного состояния водной биоты прибрежья комплекса защитных сооружений (КЗС) Санкт-Петербурга от наводнений (по материалам рыбохозяйственного мониторинга 2003-2004 гг.) // Сб. науч. тр. ГосНИОРХ. 2006. Вып. 331. Т. 2. С. 195-262.
Максимов А.А. Влияние климатических факторов на динамику макрозообентоса. Сб. Экосистема эстуария реки Невы: биологическое разнообразие и экологические проблемы / Под ред. А.Ф. Алимова, С.М. Голубкова. М.: Изд-во «Товарищество научных изданий КМК», 2008. С. 346-355.
Неелов А.В. Рыбы. Серия «Природа Ленинградской области». Л.: Лениздат, 1987. 157 с.
Никулина В.Н. Фитопланктон эстуария р. Невы в 2001-2007 гг. Сб. День Балтийского моря: тезисы докл. IX Международного экологического форума (СПб, 11-13 марта 2008 г.). СПб.: Изд-во «Диалог», 2008. С. 281-282.
Никулина В.Н., Губелит Ю.И. Водоросли литорали как показатель экологического состояния прибрежной зоны восточной части Финского залива // Биология внутренних вод. 2007. №1. С. 40-45.
Огородникова В.А., Суслопарова О.Н. Пищевые отношения массовых видов рыб Невской губы // Сб. науч. тр. ГосНИОРХ. 1986. Вып. 248. С. 126-141.
Подаруева З.С. Промыслово-биологическая характеристика невской корюшки Osmerus eperlanus (L.) по материалам 1965-1967 гг. // Изв. ГосНИОРХ. 1972. Т. 82. С. 11-35.
Попов А.Н. Питание корюшки восточной части Финского залива // Изв. ГосНИОРХ. 1978. Т. 129. С. 53-63.
Попов А.Н. О причине снижения уловов нерестовой корюшки в Финском заливе. Сб. Биологические ресурсы Белого моря и внутренних водоемов Европейского Севера: тезисы докл. II (XXV) Междунар. конф. (Петрозаводск, 22-26 ноября 1999 г.) Петрозаводск, 1999. С. 91-93.
Попов А.Н. О причине снижения численности корюшки в Финском заливе. Тезисы докл. IX Всероссийской конференции по проблемам рыбопромыслового прогнозирования. Мурманск, 2004. С. 115-117.
Попов А.Н. Биология и воспроизводство запасов корюшки (Osmerus eperlanus L.) восточной части Финского залива // Сб. науч. тр. ГосНИОРХ. 2006а. Вып. 331.
Т. 2. С. 92-118.
Попов А.Н. Многолетняя динамика состояния запасов салаки (Clupea harengus membras L.) восточной части Финского залива // Сб. науч. тр. ГосНИОРХ. 2006б.
Вып. 331. Т. 2. С. 119-139.
Попов А.Н. Воздействие экологических факторов на динамику популяций салаки (Clupea harengus membras L.) и корюшки (Osmerus eperlanus L.) восточной части Финского залива. Сб. Водные экосистемы: трофические уровни и проблемы поддержания биоразнообразия: мат-лы Всероссийской конференции с международным участием «Водные и наземные экосистемы: проблемы и перспективы исследований», Вологда, 2008. С. 343-347.
Рыбалко А.Е., Федорова Н.К. Донные отложения эстуария реки Невы и их загрязнения под влиянием антропогенных процессов. Сб. Экосистема эстуария реки Невы: биологическое разнообразие и экологические проблемы / Под ред. А.Ф. Алимова,
С.М. Голубкова. М.: Изд-во «Товарищество научных изданий КМК», 2008. С. 39-58.
Рыбалко А.Е., Федорова Н.К., Фокин Д.П., Зайцев В.М., Марковец И.М. Влияние крупных гидротехнических проектов на геоэкологическую ситуацию в Невской губе и восточной части Финского залива. Сб. День Балтийского моря: тезисы докл. X Международного экологического форума (СПб, 17-19 марта 2009 г.). СПб.: Изд-во ООО «Макси-Принт», 2009. С. 196-198.
Сендек Д.С., Студенов И.И., Шерстков В.С., Новоселов А.П., Коновалов А.Ф. Генетическая дифференциация корюшковых рыб рода Osmerus (Osmeridae, Salmoniformes) на Европейском Севере России. Сб. Лососевидные рыбы Восточной Фенноскандии. Петрозаводск, 2005. С. 148-157.
Суслопарова О.Н., Огородникова В.А., Волхонская Н.И. Воздействие повышенной мутности воды, возникающей при выполнении гидротехнических работ, на структурно-функциональные характеристики зоопланктона // Сб. науч. тр. ГосНИОРХ. 2006. Вып. 331. Т. 1. C. 274-334.
Суслопарова О.Н., Мицкевич О.И. Терешенкова Т.В., Хозяйкин А.А.,
Богданов Д.В. Особенности реакции основных компонентов биоты Невской губы Финского залива на проведение масштабных гидротехнических работ // Ученые записки Русского географического общества. Калининградское отделение. 2008. Т. 7.
Ч. 1. AZ1-AZ11, CD-ROM.
Суслопарова О.Н., Шурухин А.С., Мицкевич О.И., Терешенкова Т.В., Хозяйкин А.А. Влияние гидротехнических работ на биоту Невской губы. Сб. День Балтийского моря: тезисы докл. X Международного экологического форума (СПб, 17-19 марта 2009 г.). СПб.: Изд-во ООО «Макси-Принт», 2009. C. 206-207.
Сухачева Л.Л., Бычковка И.А., Кильдюшевский Е.И. Изучение и картирование подводных ландшафтов восточной части Финского залива на основе использования современных и архивных данных дистанционного зондирования. Сб. День Балтийского моря: тезисы докл. IX Международного экологического форума (СПб, 11-13 марта
2008 г.). СПб.: Изд-во «Диалог», 2008. С. 100-103.
Федорова Н.К., Корнеева Е.В., Ясиновская О.Ю. Результаты сравнительного анализа химического анализа придонных и поровых вод для оценки экологического состояния Финского залива в период с 2005 по 2007 гг. Сб. День Балтийского моря: тезисы докл. IX Международного экологического форума (СПб, 11-13 марта 2008 г.). СПб.: Изд-во «Диалог», 2008. С. 314-316.
Христофоров О.Л., Мурза И.Г., Медведев М.Н. Останется ли Невская губа рыбохозяйственным водоемом? Сб. День Балтийского моря: тезисы докл. X Международного экологического форума (СПб, 17-19 марта 2009 г.). СПб.: Изд-во ООО «Макси-Принт», 2009. C. 71-73.
Чичкова Е.Ф., Минаков Е.П. Мониторинг Финского залива на основе спутниковой информации. Сб. Современные проблемы дистанционного зондирования Земли из космоса. Вып. 6. 2010. В печати.
Kuhl H. Nahrungsuntersuchungen am Stint (Osmerus eperlanus L.) im Elbe-Mundungsgebiet // Arch. Fisch-Wiss. 1970. №21. Pр. 222-231.
Spilev H., Turovski A. Smelt, (Osmerus eperlanus L.). In Fishes of Estonia / Еds. by
E. Ojaveer, E. Pihu, T. Saat. Tallinn, 2003. Pр. 143-147.
Timola O. The diet of smelt, Osmerus eperlanus eperlanus (L.) in the northeastern Bothnian Bay // Bothnian Bay Rep. 1980. №2. Pр. 9-16.
THE REASONS OF DECREASE OF NEVSKAYA SMELT POPULATION© 2010 y. D.S. Sendek, А.Ye. KorolevState Research Institute on Lake and River Fisheries, Saint-PetersburgThe paper summarizes knowledge on changes of ecological situation in Nevskaya Bay (the eastern part of the Gulf of Finland) during the last several decades which had caused considerable decrease of local smelt population. Among the negative factors, which affected the collapse of the smelt, several reasons should be mentioned especially: reduction of the fish supply in the sea, the influence of the dam (of the protective constructions complex against the flood of the city), intensification of the hydro-technical works in the Nevskaya Bay, illegal fishery, not-correct estimation of the TAC. The range of the concrete measurements is suggested to improve the modern situation with Nevskaya smelt population.Key words: Nevskaya Bay smelt; fishering stock; eutrophication; hydro-technical works.
АВТОРЫ ПУБЛИКАЦИИ:
Сендек Дмитрий Сергеевич, к.б.н., ведущий научный сотрудник ГосНИОРХ
Королев Александр Евгеньевич, к.б.н., ведущий научный сотрудник ГосНИОРХ